

艾滋病毒是通过其表面膜蛋白来感染人体细胞的。这个病毒表面的跨膜蛋白是个三聚体的糖化蛋白,在病毒感染细胞的过程中构像很多,持续变化。以往在分选抗艾滋病毒的广谱中和抗体时,用来标记抗原特异性B细胞的都是游离蛋白,那么原本的病毒跨膜蛋白三聚体在失去胞膜的支撑后,往往只能以单体的形式或者固化三聚体的形式进行纯化,这些纯化的游离蛋白不太可能完全重塑病毒表面跨膜蛋白的原本构像。
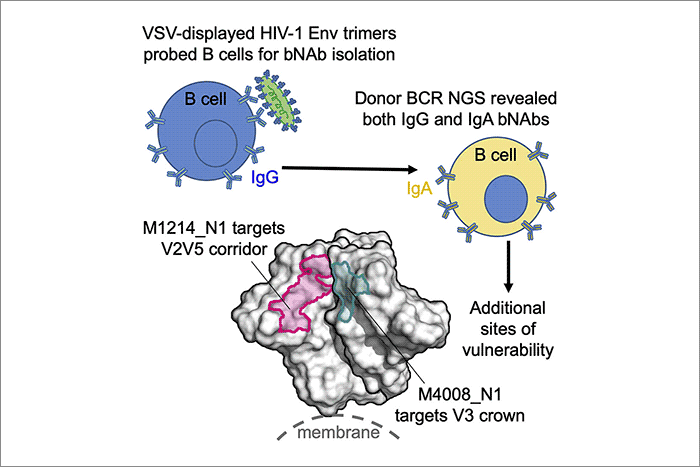
为了克服这个不足,2020年4月20日,哥伦比亚大学Aaron Diamond艾滋病研究中心伍雪玲课题组在《Cell Host & Microbe》杂志上发表文章《VSV-Displayed HIV-1 Envelope Identifies Broadly Neutralizing Antibodies Class-Switched to IgG and IgA》,首次以VSV (vesicular stomatitis virus,水泡口炎病毒)作为平台来展示艾滋病毒表面的膜蛋白,并以此来标记抗原特异性B细胞,从两个病人(代码分别为M4008和M1214)血样中总共分选出五个抗艾滋病毒的广谱中和抗体,其中三个抗体呈现出30-65%的毒株覆盖率和不错的作用强度,并在很大程度上反映了病人血浆中的总体中和抗体活性,这些结果表明用VSV展示的艾滋病毒表面膜蛋白作为B细胞标记物是成功的。
为了进一步分析这三个广谱中和抗体的B细胞谱系,课题组做了这两个病人B细胞受体(B cell receptor, BCR)的高通量测序。测序结果与这三个广谱中和IgG抗体序列做对比,不仅发现了它们在IgG同类型中的同克隆源变异体,而且对于其中的两个抗体M4008_N1和M1214_N1,还发现了IgA的同克隆源变异体!这真是意外的发现!至此为止,以往分离的抗艾滋病毒中和抗体都是IgG类型的,而这次是第一次发现IgA类型的抗艾滋病毒广谱中和抗体,而且是与众不同的转换成了IgG和IgA的广谱中和抗体!众所周知,也正如教科书所教的,抗体类型转换都是从初始IgM类型的B细胞转换成IgG或者IgA类型的B细胞。虽然有人在体外B细胞培养中或者BCR高通量测序中发现有IgG进一步转换成IgA的现象,但这种现象从未在已知功能的人源抗体中得到阐述。这个现象首次在两个病人的两个抗艾滋病毒广谱中和抗体中得到证实和明确阐述,意义重大,是对目前免疫学教科书的有益补充!
最后,课题组对这两个广谱中和抗体在病毒表面膜蛋白上的表位识别进行了分析,发现两者识别的都是新表位。M4008_N1识别的是免疫原性很强的V3顶部,而以往识别这个表位的抗体都不能有效中和艾滋病毒,所以M4008_N1识别这个表位的方式和角度一定有其独特之处。第二个抗体M1214_N1识别的则是一个从V2至V5的狭长的全新表位,因此被命名为”V2V5走廊表位”。这个抗体结合的部位是通过冷冻电镜技术取得的结构进行分析和阐述,所以十分明确、清晰。这些新表位的定义也为艾滋病毒疫苗的设计提供了新的靶位。
这篇文章的发现引人入胜。首先,课题组以VSV作为平台来展示艾滋病毒表面膜蛋白并以此来标记B细胞取得成功,克服了一个分选艾滋病毒广谱中和抗体的技术难题。其次,课题组意外发现分选出来的两个广谱中和抗体的类型已经转换为IgG和IgA,而且是通过极不寻常的从IgG进一步转换成IgA的机制。最后,这两个中和抗体识别的病毒表位都是新的。
那么问题来了,是这些新表位识别决定了这两个抗体从IgG进一步转换成IgA的命运,还是新表位识别仅仅只是巧合?IgG进一步转换成IgA的现象有多常见?仅仅是在艾滋病毒慢性感染者体内发生,还是在病毒急性感染期也会发生?别的病毒感染,包括现在全球大流行的新型冠状病毒感染,会不会引起IgG进一步转换成IgA呢?这些问题就要留待以后的抗体研究中去寻求答案了……
